猪繁殖与呼吸综合征(Porcine Reproductive and Respiratory Syndrome, PRRS)俗称“蓝耳病”,从1987年有报道以来,至今已有近30年,它仍然给全球的养猪业带来重大的损失。该病控制的困难与其致病病原“猪繁殖与呼吸综合征病毒”(Porcine Reproductive and Respiratory Syndrome Virus, PRRSV)的遗传多样性有关,随着2006年高致病性猪繁殖与呼吸综合征病毒(High Pathogenic Porcine Reproductive and Respiratory Syndrome Virus, HP-PRRSV)在我国的出现,以及后来QYYZ毒株和类NADC30毒株的报道,我国猪群存在的PRRSV毒株可能更为复杂,而在猪群意义上选择合适的措施防控该病,了解猪群中流行的PRRSV毒株则是基础。
为进一步了解我国猪群PRRSV的感染现状,2014年4月~2016年4月,我们对采自15个省56个临床存在问题的猪场的经RT-PCR检测PRRSV ORF7核酸阳性的163份样本重新进行了核酸提取,对提取的核酸进行了PRRSV ORF3、ORF5、NSP2高变区扩增、测序,通过DNAstar和MEGA6.0软件对测序结果的分析,来了解我国猪群感染PRRSV毒株的种类及其出现的频率。为减小表达的复杂性,本文仅叙述ORF5基因的调查结果和ORF3、ORF5、NSP2等3个基因综合分析得到的猪场感染毒株类型及各毒株组合的结果。
材料和方法
1.1 样品来源和分布
2014年4月~2016年4月,收集到北京、河北、山西、河南、天津、湖北、福建、广东、浙江、上海、山东、吉林、江西、四川、云南等15个省的56个保育阶段临床发病猪场的病料或血清样本,经RT-PCR检测为PRRSV ORF7核酸阳性的样本共163份,每个猪场的样本为1~5个。
1.2 主要试剂、设备、软件和方法
主要试剂及试验用溶液:病毒基因组提取试剂盒、RT-PCR相关试剂、琼脂糖凝胶电泳所用试剂及溶液、琼脂糖凝胶回收试剂盒、载体与菌株、LB液体培养基、Amp+/LB液体培养基、Amp+/LB固体培养基、SOC替代物等。
主要仪器设备:MIKRO 220R台式冷冻高速离心机、MIKRO 120台式高速离心机,德国Hettich公司产品;SW-CJ-2FD双人单面垂直净化工作台,苏州安泰空气技术有限公司产品;TW8型水浴槽,德国Julabo公司产品;2720型PCR仪,美国Applied Biosystems公司产品;DYY-11B型三恒电泳仪,北京市六一仪器厂产品;Molecular Imager Gel DocTM XR+凝胶成像分析系统,美国BID-RAD公司产品;培英HYG-A型全温摇瓶柜,江苏太仓市实验设备厂产品等。
PRRSV参考毒株核苷酸序列:本研究所用的参考毒株序列除了GDr180、HuN4-F112、TJM-F92是自行测序得到的,其它参考序列均下载于GenBank(略)。
分子生物学分析软件:DNAstar和MEGA 6.0,由本实验室提供。
方法:利用DNAstar中的Meg Align对测到的样本的ORF3、ORF5全基因和NSP2高变区基因同参考毒株的相应基因片段进行序列比对和同源性分析,利用MEGA6.0中的邻接法(neighbor-joining, boot strap values of 1000 replicates)对ORF3、ORF5全基因和NSP2高变区基因进行遗传进化分析,根据各个样本的ORF3、ORF5、NSP2比对JXA1、NADC30、VR-2332、CH-1a等毒株的结果,将其归类于不同亚群;后综合各样本3个目标基因片段的测序结果,评估各个猪场感染的PRRSV毒株类型。其它,如样品中病毒总RNA的提取、引物序列设计、引物合成、cDNA模板的制备、目的片段的PCR扩增等。
结果和分析
2.1 ORF5基因调查结果
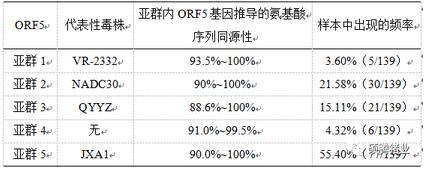
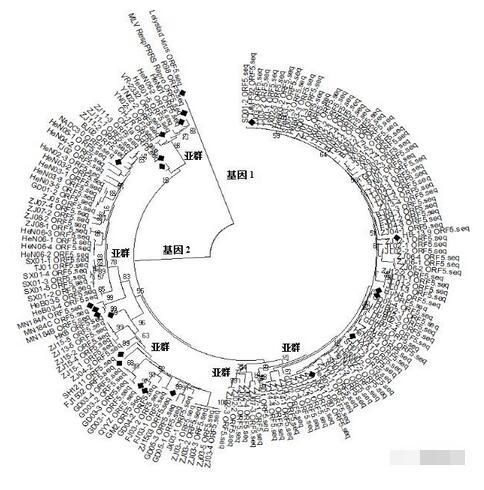
我们将得到的139个ORF5基因序列经DNAstar和MEGA6.0软件分析后发现,这些样本中的ORF5可以分为5个亚群(见图2-1),除了类VR-2332、类NADC30、类QYYZ、HP-PRRSV等4个亚群外,还发现了一个新的亚群(疑HP-PRRSV变异亚群,与CH-1a及JXA1的核苷酸同源性都在90%~92%)。各亚群代表毒株、亚群内ORF5基因推导的氨基酸序列同源性及各亚群在样本中出现的比例见表2-1。有趣的是,本研究的样本没有测到类CH-1a亚群和类经典过渡毒株。
图2-1 基于139个样本的PRRSV ORF5基因推导的氨基酸序列构建的遗传发生树
表2-1 基于ORF5的亚群划分及其在样本中出现的频率和各亚群内氨基酸序列的同源性
2.2 猪场感染PRRSV毒株调查结果
不像ORF5基因,我们在所有样本中没有测到类QYYZ毒株的ORF3、NSP2基因,但测到类经典过渡株的ORF3和NSP2基因;像ORF5一样,我们在所有样本中也没有测到类CH-1a毒株的ORF3、NSP2基因。综合分析这56个猪场的163份样本测得的130个ORF3、139个ORF5、113个NSP2基因序列的比对结果,发现来自18个猪场的38份样本的这3个基因不属于同一个亚群,而是属于2个或3个亚群。推断这3个基因测序结果不一致的样本存在的PRRSV毒株所属亚群如表2-2。
表2-2 基因测序结果不属于同一亚群的样本可能感染的毒株及其推断依据
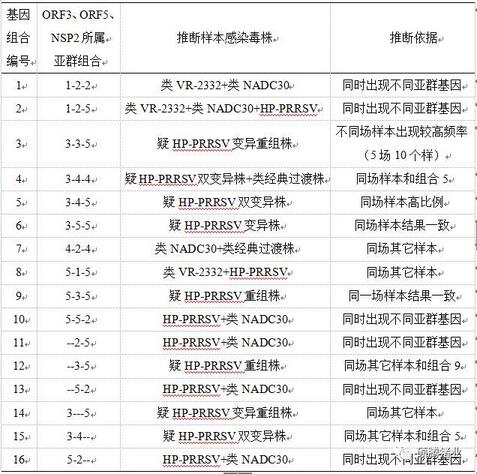
上表中的“3-3-5”、“3-4-5”、“3-5-5”、“5-3-5”因为在猪场和样本中多次出现以及NSP2基因除出现HP-PRRSV亚群基因以外无其它亚群基因,而分别推断为“疑HP-PRRSV变异重组株”、“疑HP-PRRSV双变异株”、“疑HP-PRRSV变异株”、“疑HP-PRRSV重组株”,实际上还有可能是两个或三个亚群的基因同时存在一个样本中。当然,这些疑似毒株从严格意义上讲,都应该进行进一步培养、纯化、认证。
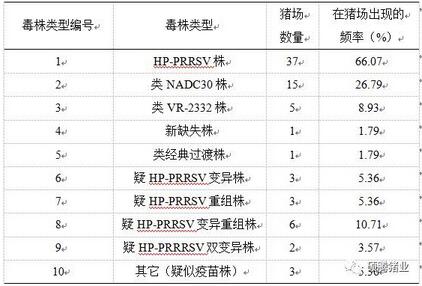
根据我们对这3个基因进行综合分析的结果,并在排除疫苗毒的可能性后(通过和各疫苗株的同源性比对),得到了目前在猪场流行的毒株类型和出现的频率如表2-3。
表2-3 各种PRRSV毒株及其在猪场出现的频率
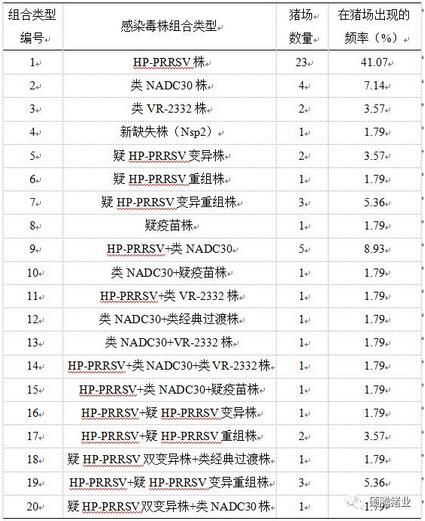
另外,我们综合分析了同一猪场不同样本的测序结果,得到各个毒株及毒株感染组合出现的频率见表2-4。
表2-4 各PRRSV毒株感染组合在猪场出现的频率
讨论
ORF5呈现复杂化
出现新的亚群。在HP-PRRSV出现以前较早的研究显示,当时我国流行的PRRSV毒株的ORF5基因可以分为两个亚群,一个较稳定,和VR-2332同源性高;另外一个亚群内同源性变化大。在HP-PRRS暴发后的2年内,有报道称当时流行的PRRSV毒株的ORF5可以分为6个亚群。稍后的报道显示,我国流行的PRRSV的ORF5可分为4个亚群,即HP-PRRSV、类CH-1a、类经典过渡毒和类VR-2332亚群。后来Xie等又研究报道了HP-PRRS发生以后的2007年~2011年华南地区PRRSV流行毒株的变化,他们把该地区这段时间流行的PRRSV毒株的ORF5分为3个亚群,即HP-PRRSV、类VR-2332、类QYYZ。Zhou等[8]在回顾当时的PRRSV流行毒株时指出,中国的流行毒株可以分为3个亚群,ORF5也可分为3个亚群,即类VR-2332、类QYYZ及HP-PRRSV。Zhang等研究分析了2014年~2015年来自东南7省的49个PRRSV毒株,他们认为ORF5可分成4个亚群,即HP-PRRSV、类VR-2332、类NADC30和新出现的亚群(经本研究分析,该亚群和QYYZ的ORF5应属同一亚群)。而由2.1部分可知,本研究样本中PRRSV ORF5基因则可以分为5个亚群,除上文提到的HP-PRRSV、类VR-2332、类NADC30、类QYYZ等4个亚群外,还发现了和经典毒株及HP-PRRSV核苷酸同源性较差的疑HP-PRRSV变异亚群(与CH-1a及JXA1的核苷酸同源性都在90%~92%)。
ORF5的HP-PRRSV亚群出现频率下降。Zhou等报道2006年~2008年我国流行的HP-PRRSV优势毒株,占88.04%。随后的报道都显示,我国流行的PRRSV,HP-PRRSV占优势地位,超过85%。而由2.1可知,由于类NADC30亚群的出现和类QYYZ亚群的增多,HP-PRRSV亚群在样本中占55.40%(77/139)。
ORF5的HP-PRRSV亚群内呈现更大的变异。Zhou等报道2006年~2008年,HP-PRRSV之间的氨基酸同源性很高,在98.3%~100%。随后的研究报道称,2009年新分离到的HP-PRRSV相对前3年的HP-PRRSV在诱骗表位和其它糖蛋白位点发生了变异,但全基因的核苷酸同源性仍很高,在98.0%~98.9%。Xie等[7]首先报道HP-PRRSV发生了较大的变化,与其代表毒株JXA1的核苷酸同源性在84.1%~99.7%。其后有研究进一步报道HP-PRRSV出现了多个分支。由2.1可知,ORF5 的HP-PRRSV 亚群和其代表性毒株JXA1的氨基酸同源性可低至93.5%,亚群内部氨基酸同源性最低为90%。
这些变化可能和以下4个方面的因素有关:1)类QYYZ和类NADC30亚群的出现。2)目前的控制措施没有很好地控制类QYYZ毒株和类NADC30毒株的传播。3)HP-PRRSV疫苗的使用降低了HP-PRRSV的传播。4)CH-1a和经典过渡毒株相对传播能力较弱,在现场已经较少,或者这两个亚群的毒株普遍毒力较弱,而我们检测的样本都是来自临床发病或问题猪场。







 增强“答卷”意识 保持“赶考”状
增强“答卷”意识 保持“赶考”状 四川省畜牧业协会20周年庆典暨2023
四川省畜牧业协会20周年庆典暨2023 华派集团组织参加四川省畜牧业协会2
华派集团组织参加四川省畜牧业协会2 解放养猪人,二厂在行动——用同孕
解放养猪人,二厂在行动——用同孕 诚聘新媒体运营专员
诚聘新媒体运营专员 脚下有泥,心中有光 | 牧原农艺师
脚下有泥,心中有光 | 牧原农艺师 携手共赢,友谊长存:江西正邦作物保
携手共赢,友谊长存:江西正邦作物保 国新办举行7月份国民经济运行情况新
国新办举行7月份国民经济运行情况新 农业农村部就2022年“三夏”生产形势
农业农村部就2022年“三夏”生产形势